Развития цнс ребенка по месяцам

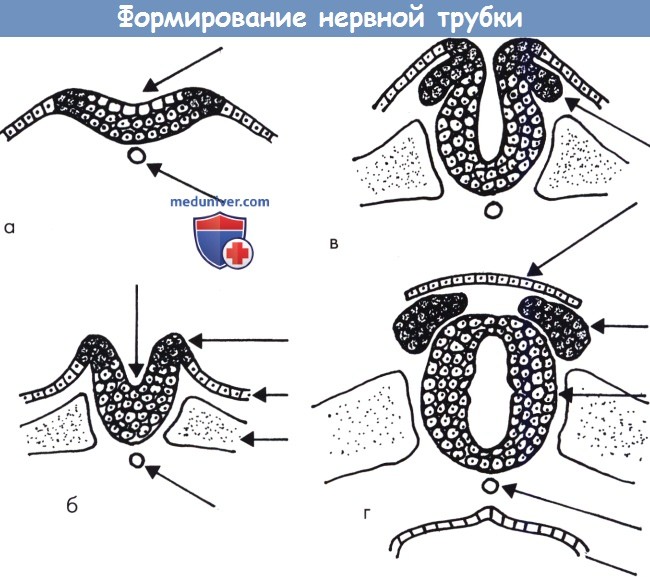
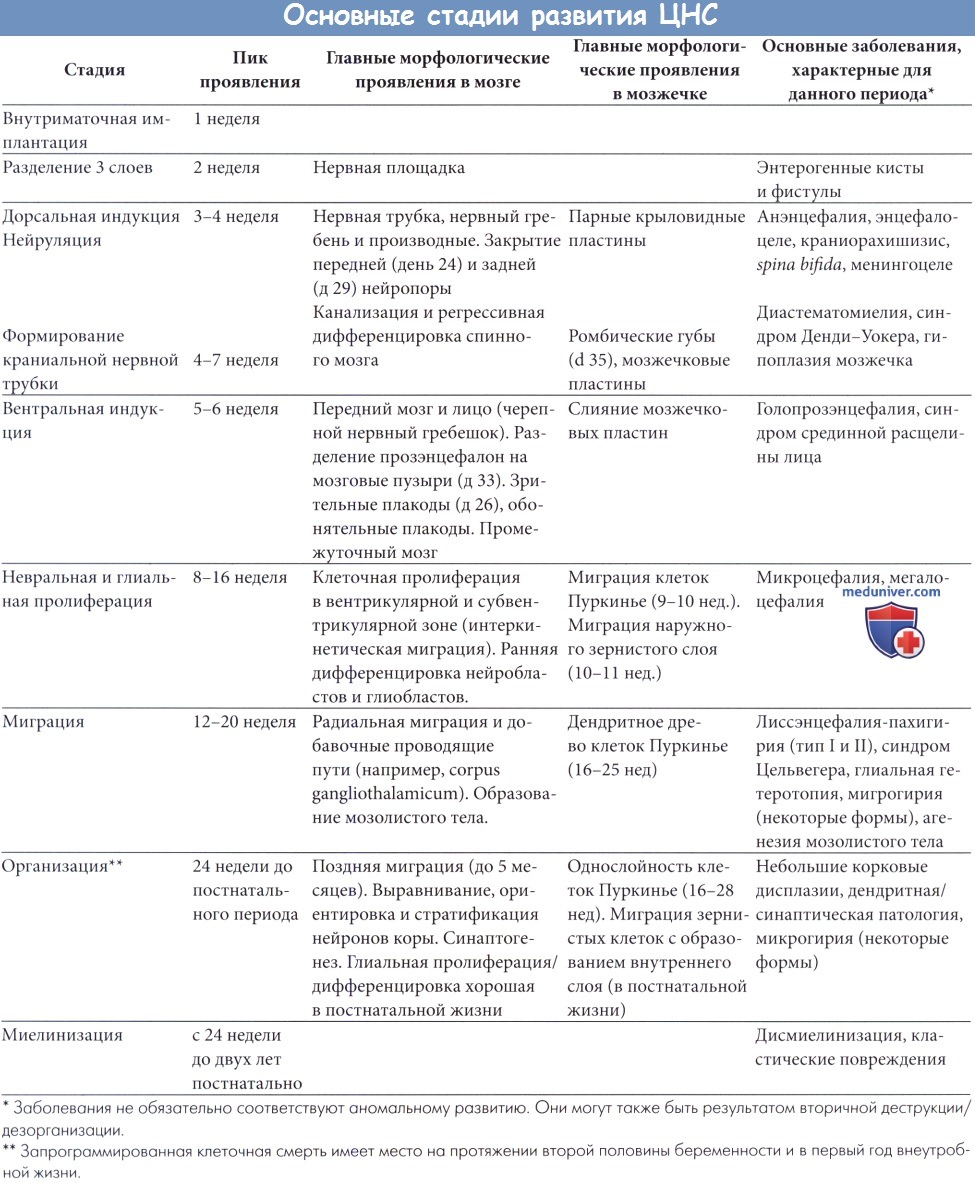
Сроки и стадии нормального развития центральной нервной системы: эмбриогенез ЦНСРассмотрим вкратце эмбриогенез ЦНС. Более полная информация содержится в классических работах (Sarnat, 1987, Barkovich et al., 1992, McConnell, 1992). В таблице ниже представлен схематический обзор основных стадий эмбриогенеза ЦНС. Регуляция развития ЦНС, фантастически сложного процесса, контролируется многими факторами, особенно генетическими. Главную роль играет большой набор белков, кодируемых или регулируемых генами (гомеобокс-гены и факторы транскрипции), которые определяют градиент дифференцировки (переднее-задний и дорсовентральный) и ограничивают определенность основных организационных единиц. Эти механизмы до сих пор в значительной степени неясны, и их изучение выходит за рамки данной книги. На протяжении второй недели эмбриогенеза формируются три слоя из эктодермы, мезодермы и энтодермы. Через две недели срединная часть эктодермы, под влиянием подлежащей мезодермы, становится нервной пластинкой, которая в дальнейшем развивается в нервный желобок, а затем в нервную трубку. На четвертой неделе нервная трубка закрывается. Процесс замыкания начинается в средней части трубки и направляется к конечностям. Молекулы адгезии нервных клеток играют центральную роль в процессе закрытия (Sarnat и Flores-Sarnat, 2002, 2004). Замыкание нервной трубки определяется дорсальной индукцией со стороны мезодермы. До закрытия трубки на девятый день на переднем конце эмбриона становятся различимыми зачатки будущего ромбэнцефалона, мезэнцефалона, прозэнцефалона и ушные плакоды. Одновременно, группы клеток вдоль боковых краев нервной трубки отделяются и формируют парные невральные гребни, которые дают начало основным структурам периферической нервной системы, оболочкам и меланоцитам. Задняя часть нервной пластины и хорда имеет различный исход: из нее формируется масса клеток, в которых в дальнейшем произойдет канализация и процесс регрессивной дифференцировки с образованием нижнего уровня спинного мозга.
К 32-33 дню в переднем мозговой пузыре формируются телэнцефалические пузыри и дифференцируется промежуточный мозг, так что на восьмой неделе представлены четыре определяемые клеточные массы в области базальных ядер. Процесс образования пузырей, по всей видимости, происходит под влиянием вентральной индукции хорды, но точный механизм индукции до конца непонятен. Приблизительно с 30 дня основные индуктивные процессы заканчиваются и начинается клеточная дифференцировка. Размножение примитивных клеток, которые затем становятся как нейронами, так и клетками глии, происходит, главным образом, около полости желудочков и, в меньшей степени, в субвентрикулярной зоне. В глубоких клеточных слоях встречаются митозы (Caviness et al., 2003). Деление клеток является асимметричным: одна дочерняя клетка мигрирует к внешней стороне во время телофазы, затем возвращается в глубокую область, чтобы начать следующий цикл; так называемая интракинетическая миграция остается в желудочковой зоне. Другая клетка покидает желудочковую зону, чтобы начать свое путешествие к кортикальной пластинке. Пролиферация нейробластов достигает максимума к 15 неделе гестации, снижаясь затем до остановки на 20 неделе. Пролиферация тесно связана с плохо понимаемым процессом запрограммированной клеточной смерти через апоптоз. Активность процессов апоптоза повышается в то же время, когда пролиферация уменьшается. Фактическая пропорция умирающих клеток у человека точно неизвестна. Вероятнее всего, этот процесс варьирует в зависимости от локализации и может в некоторых областях затрагивать более 30-50% сформировавшихся клеток. Кортикогенез частично изучен за последние два десятилетия. В позднем эмбриональном периоде (45-50 дни гестации) примитивные кортикопетальные волокна проникают через диэнцефальную борозду и распространяются под телэнцефальной мягкой мозговой оболочкой; нейроны, по всей видимости, в пределах этих волокон формируют зародышевый плексиформный слой (ИПС) (Bentivoglio et al., 2003) или препластинку. ИПС предшествует миграции нейронов, формирующей кортикальную пластинку, и служит опорой мигрирующим клеткам.
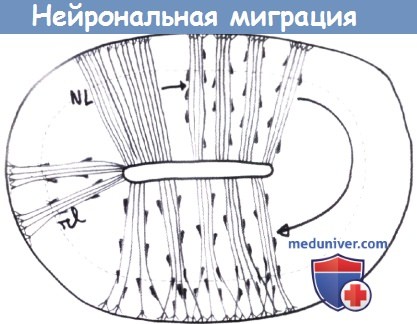
Мигрирующие нейроны делят корковую зону на поверхностную часть, которая будет составлять молекулярный слой или слой I, в основном содержащий клетки Кахаля-Ретциуса и их отростки (Sarnat и Flores-Sarnat, 2002, Rakic и Zecevic, 2003) и глубокий слой, называемый субпластинкой (или слоем VII), который исчезает до окончания внутриутробной жизни. Клетки КахаляРетциуса играют важную роль в окончательной судьбе пирамидальных клеток, вероятно, с помощью секретируемого рилина (Crino, 2001, Assadi et al, 2003). Большинство из них исчезает за счет апоптоза в возрасте до одного года. Субпластинки нейрона действуют как «переключатель» для кортикопетальных талямических аксонов до того, как клетки четвертого слоя начнут функционировать. Поздние мигрирующие клетки, которые сформируют слои со II по VI дефинитивной коры, перемещаются способом «наизнанку», то есть молодые генерированные поздне-мигрирующие клетки образуют более поверхностные слои (слой II) корковой пластины, в то время как большие пирамидные клетки слоев V и VI мигрировали раньше. Большинство нейронов будущей коры мозга перемещается между 10-й и 18-й неделями гестации, и полный комплекс корковых нейронов по существу завершается к 20-й неделе. Однако клеточная миграция продолжается в более медленном темпе на протяжении беременности, и некоторые клетки, например, зернистые мозжечковые и гиппокампальные, мигрируют постнатально (Sarnat, 1987). Некоторый корковый нейрогенез сохраняется в течение жизни, особенно в средней височной доле и мозжечке (Gage, 2002). Нейрональная миграция является сложным процессом, который отличается в зависимости от типов нейронов. Радиальная миграция затрагивает большинство (вероятно, 75%) клеток-предшественниц, предназначенных для образования пирамидных нейронов. Они мигрируют вдоль глиальных проводников, которые простираются от желудочковой (пролиферативной) зоны к пиальной поверхности нервной трубки и производных структур (Rakic, 1981, Williams и Caviness, 1984). Глиальные проводники позднее превращаются в астроциты, завершая радиальную миграцию. Несколько клеток используют одни и те же глиальные волокна при перемещении, что может отвечать за модульную организацию коры, единую для млекопитающих (Rockel et al., 1980), модули представляют собой функциональные единицы коры. Функциональные единицы видов млекопитающих отличаются количеством, а не структурой. Не все нейроны следуют точно за особым глиальным проводником. Другие клетки-предшественницы, которые затем станут интернейронами, отделяются от проводника и мигрируют в перпендикулярном направлении (Walsh, 1995, Caviness et al., 2003), часть может направиться к соседним проводникам. Некоторые клетки зарождаются в ганглионарных возвышениях в базальном мозге и мигрируют тангенциально (Jimenez et al., 2002, Bystron et al., 2005, Kanatani et al., 2005). Они следуют по до сих пор плохо известному маршруту, возможно, вдоль аксональных пучков к корковой пластине. Большинство, вероятно, интернейроны, в основном ГАМК-эргические, тогда как пирамидальные (глутаматэргические) клетки мигрируют радиально. Эти разнообразные маршруты могут объяснить широкое распространение в коре многочисленных постмиграционных нейронов, образованных из одного клона (Walsh и Серко, 1993). Мигрирующие нейробласты направляются к своим окончательным местам рядом сигнальных систем, особенно биохимическими, которые контролируются многочисленными генами, в настоящее время интенсивно изучаемыми (Sarnat и Flores-Sarnat, 2002, Crino, 2004). Вторая половина гестации характеризуется быстрым увеличением длины и сложности дендритов и аксонов посредством установления синапсов, с созреванием и тонкой организацией коры. В результате быстро увеличивается вес мозга и процесс образования борозд, необходимый для размещения интенсивно увеличивающейся площади коры. Борозды второго и третьего порядка возникают между 7-м и 9-м месяцами беременности, и большинство извилин присутствует в 28 недель. Полная ламинарная структура коры формируется к моменту рождения. Завершающим формирование центральной нервной системы моментом является снижение количества и плотности синапсов в результате развития нейрональных процессов и запрограммированной апоптозом клеточной смерти. Дендриты, аксоны и синапсы развиваются с огромной скоростью, и многие ранее образованные синусы в итоге исчезают. Развитие дендритов, аксональное ветвление и формирование дендритных корешков продолжается до четвертого года жизни. Глиальное развитие является сложным процессом, разнообразные предшественники имеют неодинаковый исход. Некоторые превращаются в радиальные глиальные волокна, используемые мигрирующими нейронами. Многие астроциты образуются из персистирующей перивентрикулярной пролиферирующей зоны после окончания процессов миграции (Gressens et al., 1992). Миелин начинает образовываться приблизительно в 30 недель, но большая часть процесса образования миелина происходит в постнатальном периоде (Yakovlev и Lecours, 1967, Brody et al., 1987).
– Также рекомендуем “Нарушения развития нервной системы: расстройства нейруляции и образования каудальной части нервной трубки (бластопатии)” Редактор: Искандер Милевски. Дата публикации: 28.11.2018 |
Источник
Нервная система ребенка. Развитие нервной системы ребенка.
Развитие нервной системы ребенка — сложный процесс, предопределяемый генетической программой развития и многообразными влияниями среды на плод и ребенка первых лет жизни.
Внутриутробное и постнатальное развитие нервной системы ребенка обусловлено постепенным, поэтапным включением отделов головного мозга и их физиологическим объединением в функциональные системы, предназначенные для обеспечения жизненно важных функций.
Поэтапность созревания заключается в постепенном переходе от филогенетически древних уровней регуляции к эволюционно более молодым и совершенным уровням, от спинально-стволового и подкоркового уровня к кортикальному. По мере этого происходит усложнение нервной системы, ее функций, возрастает степень морфо-функциональной дифференциации нервных клеток.
Системогенез в общем физиологическом смысле означает поэтапное развитие функциональных систем, обеспечивающее выживание и адекватное приспособление к условиям окружающей среды. По принципу функциональных систем осуществляется вся деятельность нервной системы.
Координация функций основана на сообщении в управляющие центры о том, что делается на периферии. Эта «обратная афферентация» обеспечивает постоянное сличение действия с первоначально заданной программой. Постоянная сигнализация о результатах действия в функциональной системе происходит в первую очередь при участии кинестезии. Кинестезии — своеобразная обратная связь, по которой корковые центры управления осведомляются о выполнении тех приказов, которые посланы на периферию.
Без этой обратной связи стало бы невозможным накопление опыта, необходимого для координации функций.
Теория системогенеза дает возможность с новых методологических позиций на основе системного подхода изучить закономерности развития функций здорового и поврежденного мозга.
В историческом аспекте условием возникновения и совершенствования специфической для человека формы деятельности и общения явилось развитие дифференцированных двигательных реакций, в первую очередь развитие руки как органа труда, а также речи, которая явилась одним из условий формирования человеческой психики.
При этом элементы функциональной системы созревают одновременно, независимо от того, к каким «горизонтальным» уровням они принадлежат.
Различие темпов формирования отдельных функциональных систем приводит к неравномерности созревания их элементов — гетерохронии роста и развития. Подобная гетерохрония наблюдается в пределах каждого «горизонтального» уровня и даже в пределах одного нервного центра. Наряду со сформированными скоплениями клеток и проводящих путей обнаруживаются морфологически и функционально незрелые структуры. У новорожденных в пределах ядра и волокон лицевого нерва в первую очередь формируются участки ядра и волокна, которые иннервируют мышцы, обеспечивающие акт сосания.
Мотонейроны VIII шейного сегмента спинного мозга гораздо раньше обеспечиваются связями со стволом головного мозга, чем соседние спинальные сегменты. Это объясняется участием двигательных клеток VIII сегмента в обеспечении рефлекса схватывания, который срабатывает уже к моменту рождения.
Концепция системогенеза существенно дополнила представления о внутриутробном и постнатальном развитии нервной системы, позволила раскрыть сущность многообразных неврологических расстройств у детей раннего возраста. Подверглось пересмотру положение о том, что новорожденный представляет собой незрелое существо, «онтогенетический нуль». В соответствии с особенностями экологии человека новорожденный ребенок «вооружен» достаточным набором функциональных систем, обеспечивающих жизненно важные физиологические акты: дыхание, сердечную деятельность, сосание и т. д. При этом отмечается отчетливая координированность этих актов. Многие функциональные системы мозга оказываются сформированными задолго до рождения и тем самым обеспечивают жизнеспособность в случае преждевременных родов.
К моменту рождения, как правило, достигается минимальное обеспечение функции, усложняющееся в процессе взаимодействия со средой. Исследование безусловных рефлексов показало, что сосательный рефлекс у новорожденного вызывается лишь при непосредственном раздражении ротовой области. В дальнейшем рецепторная зона этого рефлекса расширяется. Положение ребенка под грудью, вид бутылочки с молоком и другие условные раздражители вызывают сосательные движения, открывание рта.
Гетерохрония развития функций мозга позволяет раскрыть сущность парциальных задержек развития. В клинической практике нередко наблюдаются изолированные задержки формирования тех или иных функций. Наиболее наглядным примером являются различные варианты отставания в двигательном или речевом развитии. Могут наблюдаться моторные алалии и дислалии, дисграфии и дислексии — изолированные задержки формирования центров письма или чтения, обусловленные недоразвитием корковых центров. Встречаются кроме того случаи так называемой моторной дебильности, при которой ребенок, обладая нормальным интеллектом, не в состоянии освоить обиходные двигательные акты: застегивание пуговиц, зашнуровывание ботинок и т. д. Выявление парциальных задержек развития позволяет более целенаправленно проводить коррекционные, лечебные и педагогические мероприятия.
– Также рекомендуем “Факторы влияющие на развитие нервной системы. Окружающая среда и нервная система ребенка”
Оглавление темы “Нервная система детей.”:
1. Нервная система ребенка. Развитие нервной системы ребенка
2. Факторы влияющие на развитие нервной системы. Окружающая среда и нервная система ребенка
3. Стадийность развития нервной системы ребенка. Влияние среды на развитие нервной системы
4. Развитие речи ребенка. Развитие речедвигательного и речеслухового анализатора
5. Принципы развития нервной системы. Принципы развития речи
6. Особенности нервной системы новорожденного. Трудности оценки нервной системы детей
7. Псевдопроцессуальность нервной системы. Оценка развития нервной системы
8. Исследование нервной системы у детей. Сбор анамнеза
9. Общий осмотр в неврологии. Форма черепа
10. Окружность черепа. Выражение лица ребенка
Источник
Оглавление темы “Физиология плода. Развитие плода.”:
1. Физиология плода. Развитие плода. Предымплантационное развитие.
2. Имплантация, органогенез и плацентация.
3. Плодный (фетальный) период. Развитие нервной системы у плода.
4. Развитие эндокринной системы плода.
5. Кроветворение. Кроветворение плода. Экстрамедуллярный гемопоэз.
6. Кровообращение плода и новорожденного. Желточный период. Аллантоидное кровоообращение. Плацентарное кровообращение.
7. Сердечная деятельность плода и новорожденного. Сердце плода и новорожденного.
8. Дыхательная система плода и новорожденного.
9. Обмен веществ плода и новорожденного.
10. Выделительная система плода. Иммунная система плода.
11. Система гемостаза плода. Кислотно-основное состояние крови плода.
Плодный (фетальный) период. Развитие нервной системы у плода.
Этот период развития продолжается от 12 до 40 нед беременности.
В плодный период практически все органы и системы плода находятся в физиологическом состоянии функциональной незрелости, что и определяет своеобразие ответных реакций плода на внешние воздействия.
Согласно теории системогенеза, предложенной известным физиологом П.К.Анохиным, развитие отдельных систем у плода происходит неравномерно, при этом избирательно и ускоренно развиваются те функциональные системы, которые в первую очередь необходимы для адаптации его организма к условиям внеутробной жизни. Эта закономерность становится отчетливо заметной при рассмотрении особенностей внутриутробного развития нервной, сердечно-сосудистой, кроветворной, эндокринной и других систем.
Нервная система.
Эта система закладывается очень рано. Образование нервной трубки и мозговых пузырей отмечается уже в течение первых недель онтогенетического развития, что можно идентифицировать на эхограммах с 8_9 нед беременности, на 2-м месяце внутриутробной жизни формируются элементы рефлекторной дуги. С этого времени появляются первые двигательные рефлексы, которые хорошо заметны при ультразвуковом исследовании с 7—8 нед беременности. В дальнейшем двигательная активность плода принимает закономерный характер, в среднем составляя около трех движений за 10 мин. К 20—22 нед заканчивается период локальных проявлений рефлекторных реакций (при раздражении определенных участков тела) и появляются рефлексы более сложного характера. Например, на 21-й неделе внутриутробного развития у плода возникают первые спонтанные сосательные движения. Приблизительно в этот же период удается фиксировать появление первых электрических потенциалов мозга. К 24-й неделе двигательная активность плода внешне уже напоминает движения новорожденного ребенка.
К рефлекторным реакциям следует также отнести дыхательные движения плода. Некоторые авторы считают, что в результате дыхательных движении, которые отчетливо заметны при ультразвуковом исследовании, улучшается кровообращение плода, так как периодически возникающее понижение внурибрюшного давления способствует притоку крови в полые вены и усилению работы сердца. Дыхательные движения плода не имеют постоянного характера, они обычно сочетаются с периодами апноэ. Нельзя полностью исключить и такого предположения, что дыхательные движения плода являются подготовкой дыхательного аппарата к осуществлению его основной функции после рождения ребенка.
К концу внутриутробного периода в основном заканчивается формирование важнейших отделов центральной и периферической нервной системы плода. Однако кортикальные функции развиваются у ребенка после его рождения.
– Также рекомендуем “Развитие эндокринной системы плода.”
Источник